Onkologiya-
B.P.Kopnin
Rus Cancer Research Center. Blokhin RAMS, Moscova
sursa RosOncoWeb.Ru
01 0203 04Introducere.
Carcinogeneza - un proces în mai multe etape de mutație andother acumulare de modificari genetice care conduc la funcții de perturbații klyuchevyhkletochnyh precum reglarea proliferării și diferențierii, moartea celulelor naturale (apoptoza) reaktsiykletki morfogenetic, și probabil, de asemenea, o faktorovspetsificheskogo funcționarea ineficientă și nespecifică antitumorala immuniteta.Tolko combinație de astfel de modificări dobândite, de obicei, ca urmare a unei evoluții destul de lungă de clone neoplazice, în care există o selecție de celule cu caracteristicile dorite pot asigura dezvoltarea de tumori maligne. Veroyatnostvozniknoveniya în aceeași celulă crește multiple izmeneniyrezko genetice cu tulburari ale genomului kontroliruyuschihtselostnost sistemelor. Prin urmare, mutațiile care duc la instabilitate genetică, sunt, de asemenea, un pas esential in progresia tumorii. Bolnav, unele sisteme congenitale anomalii ale factorului kontrolyayavlyayutsya genetice determină apariția inevitabilă a unor noi formațiuni: acestea sunt atât crește probabilitatea de apariție a fiecărei kletkeorganizma diferite mutații oncogene care ransheili individual, mai târziu, în unele celule proliferante clone sub davleniemotbora se acumulează în mod necesar setul necesar de forma tumori izmeneniyi.
progrese semnificative în înțelegerea mecanismelor carcinogenezei svyazans prima deschidere și protonkogenov oncogene, și apoi - genele opuholevyhsupressorov și mutatoare. Oncogene - o celulă sau un virus (virus introdus în celula) gene a căror expresie poate privestik dezvoltarea tumorilor. Proto-oncogene - kletochnyegeny normale, îmbunătăți sau modifica funcțiile pe care le face vonkogeny. supresoare tumorale (antioncogene, opuholevyegeny recesive) - gene celulare a căror inactivare brusc apariție uvelichivaetveroyatnost a tumorilor și recuperarea funcției, prin contrast, poate suprima creșterea celulelor tumorale. Trebuie remarcat, că se face referire la așa-numitele Suprimarea tumorale "mutator"gene care Disfuncția gene care in unele ritm sposobomuvelichivaet de mutații și / sau alte geneticheskihizmeneny, nu pot afecta creșterea celulelor neoplazice. inactivare Odnakoih atât crește foarte mult probabilitatea poyavleniyarazlichnyh mutatii oncogene ca formarea tumorii stanovitsyalish o chestiune de timp.
Aparținând oncogene sau Suprimarea tumorale criterii opredelyaetsyaneskolkimi: a) caracter natural schimbă strukturyi / sau exprimarea genei în celule sau anumite razlichnyhnovoobrazovaniy- b) apariția unei tinere sau mai tinere tumori formeaza vozrasteopredelennyh la persoanele cu transmitere prin nasledstvugerminalnymi (adică a avut loc în sexual celulă) mutatsiyamidannogo gena- c) o creștere bruscă a frecvenței de apariție a opuholeyu animale transgenice care exprimă o genă activată formudannogo - în oncogenelor caz sau inaktiviruyuschiemutatsii care poartă ("knock-out") Gena - în cazul opuholevyhsupressorov- g) capacitatea de a induce în vitrokletkah cultivate în transformarea morfologică și / sau o creștere nelimitată (oncogene) sau inhibarea creșterii celulare și / sau vyrazhennostipriznakov transformare (supresoare de tumori).
Ultimele două decenii au fost caracterizate de rapidă otkrytiemvse mai mult oncogene si Suprimarea tumorii. Prin nastoyaschemuvremeni cunoscute despre sute de potențiale oncogene (virusul kletochnyhi) și aproximativ două duzini supresoare tumorale. evenimente genetice Byliopisany conducând la activarea protoonkogenovili inactivarea supresoare tumorale [1-6]. acțiune a găsit chtomehanizm oncogenelor virale asociate cu kletochnyhprotoonkogenov activare (retrovirusuri) sau inactivarea Suprimarea tumorale (virusuri ADN) [7-11]. Caracteristic pentru formele iliinyh de tumori umane și modificări opuholevyhsupressorov oncogenelor, inclusiv ispolzuemyedlya foarte specific diagnostic anomalie [3,12] (Tabel. 1, 2).
Tabelul 1.
Unele modificări caracteristice proto-oncogene de novoobrazovaniycheloveka
| protooncogen | funcţia de proteine | schimbări | neoplasme * |
|---|---|---|---|
| RET (GDNF-R) | receptor tirozin kinazei | a) activarea mutanților ghiduri de puncte; b) recombinare ob formatoare de gene himere Ret / ptc, codificării de fierbere-permanent ACTI-receptor virovanny | sindroamele endocrine multiple neop-leneș (MEN2a, MEN2b), medular și cancere papilare * tiroidian |
| ErbB1 (EGF-R) | receptor tirozin kinazei | Amplificarea și supraexprimarea genei | glioblastom și alte tumori neurogene |
| ERBB2 (HER2) | receptor tirozin kinazei | Amplificarea și / sau supraexpresia genei | cancerul mamar |
| PDGF-Rb | receptor tirozin kinazei | cromozom-translațională katsii formeaza gene chi-dimensionale TEL / PDGF-Rb, CVE6 / PDGF-Rb, Cody constant ACTi-al-receptorii virovannye | leucemie mielomonocitară cronică, leucemia mieloblastică acută |
| SRC | non-receptor tirozin kinazei | mutații la nivelul codonului 531, desființând reglarea negativă a activității kinazei | tumori de colon parte în stadiile tardive |
| K-RAS, N-RAS, H-RAS | implicată în transmiterea semnalelor mitogene și a reacțiilor-morfogeneti cal de reglare | mutatii in codonii 12,13,61, provocând formarea de formă legată de GTP-activat permanent Ras | 60-80% din cazurile de cancer pancreatic zhelezy- 25-30% din diferite tumori solide și leucemii |
| PRAD1 / tsiklinD1 | regleaza ciclul adeziv precis | Amplificarea și / sau supraexpresia genei | cancer de san si glandele salivare |
| C-myc | Transkom factor-RTOC, regulatori ai ciclului liruet-kletoch-lea și activitatea telomerazei | a) trans-cromozom localizare, transmițând gena sub controlul elementelor de reglare ale genei de reglementare imunoglobulină; b) amplificarea și / sau supraexpresia gena- mutație stabilizează proteina-ing | a) limfom Berkita b) mai multe forme de tumori |
| CTNNB1 (b-catenin) | a) factor de transcriere-onny, reguliret c-myc și cyclin D1; b) legarea la caderină predare există în contactele adezive guvernamentale mod Vania | mutație care crește cantitatea de nelegată conductor legat la E-caderinei b-catenin, care funcționează ca un factor de transcriere | polipoza adenomatoasă familială colon kishki- diverse forme de tumori sporadice |
| BCL2 | inhibă apo ptoza, reglarea permeabilității mitocondrie si membrane Nucleii IAL-TION | cromozom-translațională katsii, transmițând gena sub controlul elementelor de reglare ale genei de reglementare imunoglobulină | limfom folicular |
| ABL | regleaza ciclul celular si apoptoza | katsii-cromozom de translație care conduce la Obra-mations gene himere BCR / ABL, toryh-produse pentru a stimula proliferarea celulară și apoptoza podvlyayut | toate leucemie mieloidă cronică, leucemie limfatică acută parte blastic |
| MDM2 | inactivează p53 si PRB | Amplificarea și / sau supraexpresia genei | o parte din osteosarcom și sarcoame ale țesuturilor moi |
Tabelul 2.
Forme de tumori umane, care rezultă din Suprimarea inactivarea genelor nekotoryhopuholevyh și mutatoare
| genă | funcţia de proteine | neoplasme * |
|---|---|---|
| p53 | regulatoare factor de transcriere-liruet ale ciclului celular și apoptoza, monitorizează integritatea genomului | Sindromul Li-Fraumeni și cele mai multe forme de tumori sporadice |
| INK4a-ARF | inhibarea Cdk4 **, activarea p53 ** | melanomul ereditar si multe tumori sporadice |
| rb | Controalele intrare în fază S, reglarea activității factorului de transcripție E2F | retinoblastom ereditar si multe forme de tumori sporadice |
| TBR-II | receptor pentru al doilea tip de citokine TGF-b | cancere ereditare si sporadice ale colonului |
| Smad2, SMAD 3 | transmiterea unui semnal de activat receptorii TGF-b la Smad4 | cancerul de colon, plămân, pancreas |
| SMAD4 / DPC4 | transcriere efect factor-OPOS-reduet a citokinei TGF-b, ceea ce conduce la activarea ingibito-șanț Cdk - p21WAF1, p27Kip1, p15INK4B | polipoza juvenilă gamartomatoz-lea stomac kishechnika- și diverse forme de tumori sporadice |
| E-cadherin | implicate în interacțiunile celulă-celulă, deystviyah- inițiază transmisia semnalelor care activeaza p53, p27Kip1 | cancere de stomac ereditare și multe forme de tumori sporadice |
| APC | distruge și se leagă tsitoplaz nematic-b-cateninei, previne formarea de complexe a-transcripțional b-catenin / TCF | polipoză familială adenom-tozny si tumorile de colon sporadice |
| VHL | inhibă expresia genei VEGF (factorul de creștere endotelial vascular) și alte gene care sunt activate in timpul hipoxie | sindromul von Hippel-Lindau (hemangioame multiple) - carcinom cu celule renale cu celule clare |
| WT1 | factor de transcriere de legare la p53 modulează genele de elaborare a politicilor Ex-Press p53-receptiv | nefroblas ereditare toms (tumora Wilms) |
| PTEN / MMAC1 | fosfataza- promovează apoptoza prin inhibarea activității PI3K-PKB / Akt de semnalizare | Boala Cowden (The hamartoame set-guvernamentale) - multe tumori sporadice |
| NF1 (neyrofibromin) | proteine de familie GAP-ras oncogen se transformă dintr-un activ într-o formă inactivă | neurofibromatoza primul tip |
| NF2 (Merlin) | implicate în interacțiunea cu citoscheletul membranei | neurofibromatoză doilea tip meningita sporadice Ohmi, mezoteliom și colab. Tumora |
| BRCA1 | crește activitatea p53 și alți factori de transcripție, legarea la RAD51 participă la învățare și / sau repararea ADN-ului | tumorile de san ereditare si diverse forme yaichnikov- tumori sporadice |
| BRCA2 | factor traskriptsionny cu activitățile acetil histone transferazy- legarea la RAD51 este implicată în repararea ADN-ului | tumorile de san ereditare si diverse forme yaichnikov- tumori sporadice |
| MSH2, MLH1, PMS1, PMS2 | Site-uri nepereche repararea ADN-ului (reparații asimetrie) | nonpolyposis cancer de colon si yaichnikov- multe tumori sporadice |
** locus INK4a / ARF codifică două proteine: p16INK4a - inhibitor tsiklinzavisimyhkinaz CDK4 / 6 și p19ARF (Alternative Frame Reading) - produs de lectură alternativnoyramki care legarea p53 si Mdm2, blocând interacțiunea lor previne degradarea p53 [13, 14]. Ștergerile și multe tochechnyemutatsii locus INK4a / ARF supressornyhaktivnostey determina inactivarea simultana a ambelor proteine [15].
Cu toate acestea, cunoștințele de mult timp despre fiecare dintre oncogene sau opuholevyhsupressorov prezentate în discret, în mare parte fără legătură. Și numai în ultimii ani a devenit imagine vyrisovyvatsyaobschaya, care arată că marea majoritate izvestnyhprotoonkogenov si tumorale Suprimarea sunt komponentamineskolkih cai comune de semnalizare care controleaza ciclul celular, apoptoza, integritatea genomului, și differentsirovkukletok reacția morfogenetice. Evident, schimbările în aceste căi de semnalizare în kontsekontsov și să conducă la dezvoltarea novoobrazovaniy.Dalee maligne oferă informații cu privire la principalele obiective ale acțiunii onkogenovi Suprimarea tumorale.
1. oncogenelor și Suprimarea tumorale în reglarea ciclului celularBaza de formare a tumorii este excesivă opredelennyhkletok reproducere. Este firesc, prin urmare, că încălcările regulyatsiikletochnogo ciclu este un celule integrale și fundamentale priznakomneoplasticheskoy. "motor" ciclului celular, așa cum se știe, sunt secvențial activitatea drugdruga succesivă a kinazelor dependente de ciclină [16] (fig. 1). Fiecare tsiklinzavisimayakinaza (Sdk) este catalitică holofermentnogokompleksa subunitatea pentru care activitatea necesită aktivatornoysubedinitsy prezență - ciclină. Regulament al scorului activității nivelul regia Sdk osuschestvlyaetsyaza schimba anumite cicline vopredelennye fază a ciclului celular. Mai mult, activitatea schimbă Sdkreguliruetsya fosforilarea anumitor aminokislotnyhostatkov. Forma activă a complexelor ciclină-CDK fosforiliruyutregulyatornye proteine care controlează curgerea fazei.

Fig. 1. Mișcarea ciclului celular este determinat cyclin posledovatelnoyaktivatsiey diferite - Cdk. Cele mai multe dintre ele este tinta efectul de activare inhibarea oncogene sau deystviyaopuholevyh supresoare
Sa constatat că efectul multor proto-oncogenelor și tumora supressorovnapravleno reglarea diferitelor complecși ciclin - produse Cdk.Belkovye cele mai multe dintre ele cresc activitatea tsiklinzavisimyhkinaz responsabilă pentru etapele inițiale presynthetic fazyG1 (complexe de cicline D1 - D3 c Sdk4 sau Sdk6 funcție ottipa celule) tranziția de la faza G1 în sinteza ADN-ului (ciclina E - Sdk2) (Figura 1.). Mai mult, unele proto-oncogenelor și activitatea supressoryreguliruyut tumorală ciclinei A - Sdk2 (dlyareplikatsii ADN necesar) și ciclinei B - Cdk1 (de asemenea, cunoscut sub numele de Cdk1 - Cdc2, este necesară pentru tranziția de la G2 în mitoză).
Substratul primar al complecșilor ciclin D - Cdk4 și ciclină D - Cdk6yavlyaetsya supresoare tumorale pRb si proteine Rb-like și p105 r130.pRb defosforilat și omologii săi în celulele care nu se divid si celule vproliferiruyuschih sunt în G1-fază timpurie [17]. VTAK starea ei se leaga si blocul de transcriere kompleksyE2F - DP (E2F-1, -2, -3, -4, -5, și DP-1, -2, -3), aktivnostryada reglarea genelor ale căror produse sunt necesare pentru începutul și prohozhdeniyaS- faza. În particular, E2F-DP reglementa expresia genelor a timidin kinazei, dihidrofolat reductaza, ciclina E, ciclină A, PCNA, DNA polimerazya și colab. [18]. Legarea proteinelor familiei E2F c PRB ingibiruetih activitatea transcripțională. Când semnale mitogene vyzyvaemyhrostovymi factori, pRb la mijlocul lui G1-faze kompleksomtsiklin fosforilată D - Sdk4 (sau ciclina D - Cdk6), care determină factorii E2F vysvobozhdenietranskriptsionnyh - DP din complexul cu pRb și activarea acestuia [17]. O consecință a acestui fapt este stimularea transkriptsiigena ciclina E, prin complexele activate ale ciclină - CDK2, de asemenea, fosforilează pRb. Astfel voznikaetregulyatornaya balama activitate de susținere transkriptsionnyhfaktorov E2F - DP si gene controlate de a furniza replikatsiyuDNK (a se vedea figura 2..). După terminarea fazei S devine pRb defosforilirovannoesostoyanie în care blochează activitatea E2F - PD și după intrarea fazei S (pentru inițierea sa necesită o nouă mitogennyystimul activarea complexelor ciclină D - Cdk4,6). Astfel, supresoare tumora PRB joaca un rol-cheie în vhozhdeniyakletki de reglementare în S-fază.
Produsele multor proto-oncogene sunt componente signalnyhputey responsabile pentru activarea ciclinei D - Cdk4 (6) și ciclina E - Cdk2 ca răspuns la factorii de creștere și / iliadgeziyu celule la proteinele matricei extracelulare (figura 2.). Astfel, legarea receptorilor factorului de creștere cu liganzii lor indutsiruetdimerizatsiyu autofosforilare și receptor (un subedinitsadimera phosphorylates alt tirozina). Aceasta, la rândul său, determină interacțiunea receptorilor tirozin kinaze cu multe signalnymibelkami conținând domenii SH2-legare, fosfotirozină. De exemplu, activat receptorii factorului de creștere al trombocitelor (PDGF-Rb) interacționează cu SH2-domenii de proteine, cum ar fi fosfatidilinozitol-3`-kinazei (PI3K), fosfolipaza C (PLC) -g1, formele latente și proteina transkriptsionnyhfaktorov STAT adaptor Grb2, transmiterea semnal la belkamRas [19-21]. Legarea fiecare dintre aceste proteine cu suprapunere fosfotirozinamiretseptora provoacă activarea căilor de semnalizare și sunt urmate de activarea in nucleul de expresie transcriere kit faktorovi genelor specifice (Fig. 2). În particular, indutsiruemyyGrb2 tranziția la o proteină activat Ras sostoyanievedet (legat-GTP) pentru a stimula unele efectorilor sale, inclusiv serina treoninovyhkinaz Raf * și MEKK, declanșând MAP (Mitogen Activated Protein) cascadele kinază [20,22]. Produsele finale ale acestor cascade, ERK (MAPK), p38 și JNK (SAPK), translocate din citoplasmă la nucleu unde fosforilează și activează multe substraturi, tomchisle astfel de factori de transcripție cum ar fi, Elk1, Ets1 *, Ets2 *, iunie *, ATF2 , TCF și altele. Aceasta, la rândul său, determină aktivatsiyuryada alți factori de transcripție. Astfel, Elk1, formând komplekss SRF (Serum Factor Response), inițiază transcripția genelor care conțin în elementele sale promotor EOA, cum ar fi FOS * gena.

Fig. 2. Produsele multor proto-oncogenelor și activitatea supressorovreguliruyut tumorală a kinazelor dependente de ciclină fosforilare pRb.Fosforilirovanie pRb, precum și legarea sa la următoarele oncoproteins virale, provocând eliberarea și activarea transcripției kompleksovE2F - DP, expresia creșterea genelor ale căror produse neobhodimydlya trecerea fazei S
Reacții similare sunt observate la integrina legare (receptor care mediază adeziunea celulară) cu proteine ale interacțiunii matriksa.Takoe extracelular determină activarea și autofosforilirovaniekinazy FAK (Focal Adhesion Kinase), prin care svyazyvaetsyas domeniu SH2 protoonkobelka Src, care la rândul său vyzyvaetrekrutirovanie proteină adaptor Grb2, activarea Ras și kinaznyhkaskadov MAP (Figura 2).
Consecința schimbărilor în activitatea unui număr de factori de transcripție induși activarea MAP kinazelor este D1 creștere ekspressiigena ciclina (se presupune că este responsabil pentru belkiJun, Ets1, Ets2 [21]). Mai mult, semnalele mitogene povyshayutekspressiyu Myc, ceea ce determină o creștere a activității tsiklinzavisimyhkinaz operează în G1 (ciclină D - Cdk4 și ciclina E - Cdk2). Etosvyazano că myc, în primul rând, transactivează gena Cdc25a- fosfatazei care elimină fosforilarea de inhibare a CDK2 și Cdk4po Thr-14 și Tyr-15, iar în al doilea rând scade expresia ingibitoraSdk2, p27KIP1a [23-26]. mecanismele de activare Myc sub influența rostovyhfaktorov studiat până rău. Se presupune că acesta poate privoditkak cai de semnalizare Ras-independente activate oncoprotein Src sau cascadele care cauzează activarea Ets1i / sau E2F Ras-Raf-MAP-kinaza (gena MYC cuprinde elemente care răspund un promotor dlyaetih factori de transcripție [22,26]) .
Mulți participanți la căile de semnalizare care mediază deystvierostovyh ca răspuns la activarea factorilor kinaze dependente de ciclină și, prin urmare, stimularea diviziunii celulare, sunt proto-oncogene. Structura Izmeneniyaih (mutație), care rezultă din evadarea factorilor de reglementare vozdeystviyanegativnyh și / sau povysheniyuekspressii permanent convertit protooncogene la oncogene astfel [1,3,5] .Cele produsul identificat oncogene sunt semnal toate etajele regulyatsiimitogennogo [5]: factori de creștere - PDGF-b (Sis), FGF1i alții-receptor tirozin kinaze - EGF-R (ErbB), HGF-R (Met), Ret și proteine altele familie Ras - K-Ras, H-Ras și N-Ras- effektoryRas - serina treonin kinaze Raf și factori-Jun MOS de transcriere, Ets1, myc, etc și, în final, ciclina D1 (PRAD1). Skladyvaetsyavpechatlenie că o analiză detaliată a fiecărui novoobrazovaniivyyavlyayutsya modificări cel puțin una dintre componentele signalnyhputey (protooncogene) determinând stimularea permanentă aktivnostitsiklinzavisimyh kinazei și inițierea diviziunii celulare este acțiunea zavisimostiot de factori de creștere.
Interesant, calea de semnalizare Cdk-Rb-E2F este controlată nu tolkopRB, dar, de asemenea, multe alte proteine supresoare (Fig. 2). Nekotoryeiz ele sunt subunități inhibitoare de Cdk (CKIs - Inhibitori de Cdk), mediatoare ciclului celular de arestare ca răspuns la razlichnyevnekletochnye și a semnalelor intracelulare [16]. Familia Identifitsirovanodva CKIs: INK4 și Cip / Kip. Primul este format din patru membri, inclusiv Suprimarea tumora p15INK4B și p16INK4a. Proteinele Ink4obladayut specificitate suficient de îngust: asociindu Cdk4 și Cdk6, previn formarea complexelor lor cu cicline D [16,27] .Semeystvo Cip / Kip este compus din trei membri: p21WAF1 / CIP1, p27KIP1ai p57KIP2. Aceste proteine se leagă la și inhibă ciclina deja complet sformirovannyekompleksy D - Cdk4 (6), ciclina E - CDK2 și ciclină A - Cdk2.Krome plus, p21WAF1 / CIP1 este capabil să blocheze și tsiklinB complexe - Cdc2, este responsabil pentru promovarea G2 faze și intrarea mitozei [16,27]. Și p21WAF1 / CIP1 și p27KIP1a mediază influența drugihsupressornyh proteine. p21WAF1 / CIP1 este un important p53 acțiune misheneytransaktivatsionnogo, și în consecință supresor implicat în reglarea activității de stabilitate / p53 (p19ARF, ATM, WT1 [13,14,28,29] sau activitatea de transcripție (BRCA1 ip33ING1 [30-32 ]). (BRCA1 și WT1 capabil să activeze p21WAF1 / CIP1takzhe si mecanisme p53 independente încă necunoscute [31,33]).
Împreună cu r15INK4b, p27KIP1a este o componentă cheie peredachiingibitornyh semnalele induse prin legarea TGF-b cu svoimiretseptorami (Fig. 2). Recent a constatat că activarea retseptoryTGF-b fosforila specifice efectorilor de semnalizare belkiSmad2 și Smad3, determinând legarea lor la tumora supressoromSmad4. Complexele rezultate vyadro transloca din citoplasmă, unde reglează transcripția genelor specifice in chastnostiingibitorov Cdk. Ca rezultat, activat și p21WAF1 / CIP1 și r15INK4b [34-37]. Ultima p27KIP1a dislocă din complexul cu Cdk4 / 6 Formarea ipodavlyaet a complexelor lor cu cicline D, neobhodimyhdlya avansarea G1 (Fig. 1). P27KIP1a eliberată este, în leaga svoyuochered și inhibă complexele ciclinei E - Sdk2, începutul otvetstvennyeza fazei S. Expresia crescută a p21WAF1 / CIP1 vedetk suprimă de asemenea activitatea ciclinei D - Cdk4,6 și tsiklinE - CDK2. Ca rezultat, celula se oprește în G0 / G1 și S faze vhoditv (Fig. 3).

Fig. 3. Legarea TGF-b la receptorul său determină complexele obrazovanietranskriptsionnyh Smad4 - Smad2,3, care translotsiruyutsyaiz citoplasmă la nucleu. Aceasta conduce la activarea unui număr de ținte, inclusiv greșit și ciclină-dependente inhibitor al kinazei p21WAF1 / CIP1, p15INK4B, p27KIP1a, care determină activitatea supresie Cdk4,6 și CDK2, otvetstvennyhza G1 avansarea și intrarea în fază S (explicații în text)
Efectul inhibitor al TGF-b sau depăși onkogenovMYC supraexpresia MDM2 [38]. Și dacă acțiunea asociată cu activarea myc razlichnyhCdk prin creșterea expresiei cdc25A [23] și de stimulare degradatsiip27KIP1a [24], proteina Mdm2 în plus degradarea p53 [13,14] inactivarea vyzyvaeti a pRb [39], eliberând astfel E2F transkriptsionnyekompleksy activ - DP. În consecință, supraexpresia protoonkogenovMYC sau MDM2 și inactivare mutații în tumoră supressorahSmad4, p15INK4B, pRb au o consecință comun - celule uskolzayutot efect inhibitor TGF-b, care apare foarte vazhnymdlya tumorilor epiteliale, în special cancerele pancreatice kishechnikai [40,41] .
Una dintre cele mai remarcabile realizări din ultimii doi ani yavilasidentifikatsiya alta cale importanta de semnalizare, chastonarushayuschegosya foarte intr-o varietate de tumori umane și desfășoară este, probabil, reglarea ciclului celular, în funcție de structurile și celulele [42] (vezi. Fig. 4) .Okazalos sostoyaniyamembrannyh submembrane, care nu are legătură cu starea Ecaderinei functiei b-kateninmozhet ca un factor de transcriere. Citoplasma onsvyazyvaetsya cu un alt factor de transcriere Tcf4, după chegokompleksy b-cateninei - Tcf4 transloca nucleu și aktiviruyutgeny având în componența sa specifică exercitarea receptiv elementy.Odnimi obiective majore de acțiune transactivation kompleksab-catenin - Tcf4 este genele cyclin D1 [43] și MYC [44]. Opuholevyysupressor APC, mutatii congenitale in care determina dezvoltarea adenomatoznogopolipoza intestinului se leaga citoplasmatice liber b-catenin, care provoacă degradarea acestora din urmă [45,46]. Astfel, stimulând formarea complexelor inaktivatsiyaARS b-catenin - Tcf4, povyshaettranskriptsiyu gena cyclin D1, myc și ca o consecință, conduce la aktivatsiitsiklinzavisimyh kinazele responsabile G1 promovarea vhodv și faza S (Figura 4.). Același efect conduce mutatsiib-catenin, care crește stabilitatea în citoplasmă (takiemutatsii gasite la pacientii cu polipoza familiala, nu APC imeyuschihmutatsy [43,47] - cele mai multe dintre aceste site-uri afecteaza b-cateninei, glicogen sintetaza fosforilat kinază-3b, GSK -3b). Shodnayakartina observate, de asemenea, atunci când oncogen activat WNT1 (Wingless / INT1) (Figura 4). Legarea acesta produsul Wnt1 (un membru al familiei cisteinei bogatyhglikozilirovannyh proteine de semnalizare) la receptorul său (carliontat) cauzează citoplasmic proteina DSH la membrana unde inhibă activitatea kinazei GSK-3b, care fosforiliruyab-catenin și APS, stimulează legarea lor și degradarea b mod -katenina.Takim Wnt1 inactivarea indusă de GSK-3bprivodit pentru a stabiliza și a crește kontsetratsiitsitoplazmaticheskogo intracelular b-catenin, care crește probabilitatea complexelor de transcripție obrazovaniyaaktivnyh s b-cateninei cu factori semeystvaTcf / Lef1 [48]. Este posibil ca mutatii in Ecaderinei takzhemogut este responsabil pentru stimularea căilor de semnalizare ale activităților oposreduemyhtranskriptsionnymi b-catenin.

Fig. 4. Mutatii tumorale APC supresoare și b-cateninei, kaki activarea oncogenelor wnt1, stimulează formarea transkriptsionnyhkompleksov b-catenin - Tcf4, gene de reglementare myc și D1.V ciclinei rezultatul creșterii activității unei serii de cyclin - Cdk (explicații în text)
Pentru a rezuma această secțiune, observăm că cele mai multe izvestnyhprotoonkogenov si tumorale Suprimarea in orice mod kinaze reguliruyutaktivnost ciclină dependente responsabile de intrarea în ciclul S-fazukletochnogo. Produsele unora dintre celule (Mdm2) sau virale (T SV40 virus antigen, adenovirus E1A, E7 HPV și colab.) Și onkogenovsvyazyvayut principal substrat inactiva astfel de Cdk - PRB. Se pare că, în încălcarea căilor de semnalizare > Cdk2,4 / 6 > pRB > E2F / DPyavlyayutsya necesare pentru apariția celulelor în mod constant proliferiruyuschihneoplasticheskih.
 Vârsta copilului în dezvoltarea tumorilor. Efectele prenatale asupra carcinogeneza
Vârsta copilului în dezvoltarea tumorilor. Efectele prenatale asupra carcinogeneza Controla diferențierea celulelor stem. Accelerarea proliferării celulelor stem
Controla diferențierea celulelor stem. Accelerarea proliferării celulelor stem Terapia cu celule stem nu duce la cancer
Terapia cu celule stem nu duce la cancer- Oamenii de știință au descoperit o cauza genetica de leucemie limfoblastică acută
- Oamenii de știință au descoperit o gena care leaga cancerul de san si tiroidian
- În produsele vulnerabile genetic de oameni din carne duce la cancer colorectal
- Eterogenitate intratumorală determină rezultatul tratamentului cancerului
- Onkologiya-
- Onkologiya-
- Onkologiya-
- Onkologiya-
- Onkologiya-
- Onkologiya-
- Onkologiya-
 Onkologiya-
Onkologiya- Onkologiya-
Onkologiya-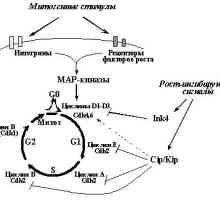 Onkologiya-
Onkologiya-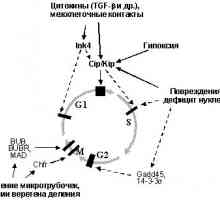 Onkologiya-
Onkologiya-- Onkologiya-
- Onkologiya-
- Onkologiya-
 Genetica cancerului
Genetica cancerului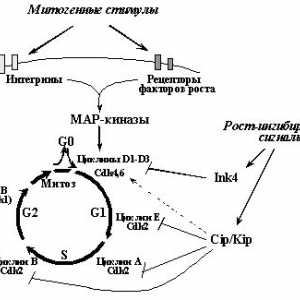 Onkologiya-
Onkologiya- Onkologiya-
Onkologiya-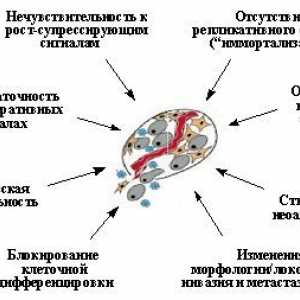 Onkologiya-
Onkologiya- Vârsta copilului în dezvoltarea tumorilor. Efectele prenatale asupra carcinogeneza
Vârsta copilului în dezvoltarea tumorilor. Efectele prenatale asupra carcinogeneza