Onkologiya-
B.P.Kopnin
Rus Cancer Research Center. Blokhin RAMS, Moscova
sursa RosOncoWeb.Ru
01 0203 0405 06073.4. produse de gena INK4a - p16INK4a si PARF - reglementa p53 si aktivnostpRb
P53 următor după schimbările de frecvență din diferite gene novoobrazovaniyahcheloveka INK4a este situat în scurt hromosomy9 braț (segment 9p21). Caracteristica sa excepțională este odnovremennoekodirovanie doua proteine nonhomologous nucleare - p16INK4a si PARF (produse de cadre alternative de citire), fiecare funcției supresoare kotoryhvypolnyaet (Figura 5.).

Fig. 5. gena INK4a codifică două non-omoloage funcție proteină vypolnyayuschihraznye.
p16INK4a leagă kinaza dependentă de ciclină complecși activi funcțional Cdk4 și cdk6 și prepyatstvuetobrazovaniyu cu cicline D, care fosforilare pRb, care inițiază intrarea în faza S. kletochnogotsikla. PARF are capacitatea de a stabiliza și p53 aktivirovatbelok, perturba interacțiunea cu proteina Mdm2 (vezi. razdel3.3). În plus, el a identificat și funcțiile-p53 independente. Astfel, se poate, de asemenea, interacționa direct cu mishenyupRb de bază - proteina E2F si bloca activitatea. Mai mult decât atât, stabilitatea onponizhaet a factorului de transcripție HIF-1 (vezi fig. 3.3.2i 3.5.3), asociind-o subunitate.
Activitatea și PARF p16INK4a creșteri în exprimarea unui număr de viral (E1A) și celulară (RAS, RAF, myc și colab.). Oncogenele O astfel de element de receptiv prezența effektobuslovlen genei INK4a dlyatranskriptsionnogo factor E2F, activabil multe mod onkogenami.Takim, produsul genei normale INK4aeffektivno operație previne multiplicarea celulară ulterioară în activarea kotoryhproizoshla orice reprezentanți ai unui gruppyonkogenov mare. În plus, expresia p16INK4a crește cu contacte obrazovaniimezhkletochnyh care asigura contactul tormozhenierazmnozheniya celulele.
șoarecii transgenici knockout homozigoți expresia genelor INK4a vyzyvayuschimpoteryu atât a produselor sale proteice observate apariției vysokayachastota în vârstă tânără de diferite neoplasme, limfoame și preferabil fibrosarcoame (ca șoarecii inaktivirovannymr53). Este interesant faptul că aproape aceeași imagine nablyudaetsyai soareci gena INK4a rearanjare, având ca rezultat pierderea de proteine ekspressiitolko PARF. Dimpotrivă, șoarecii au pierdut ekspressiyup16INK4a dar păstrat expresia PARF, nu povysheniyachastoty semnificativă a tumorilor (indicat doar vozniknoveniemelanom rare nu au fost observate la tulpinile de șoarece parentale). In vitro, fibroblaste murine nu kulturahin ekspessiruyuschie genaINK4a ambele produse, precum și fibroblaste, în care numai inactivat PARF, sunt ușor transformate RAS familie oncogene. La transformarea celulară același vremyadlya Ras indusă, nu ekspressiruyuschihtolko p16INK4a, necesită acțiuni suplimentare, cum ar fi kooperiruyuschihonkogenov MYC sau E1, la fel cum este cazul nablyudaetsyav celulele normale.
Oameni mutatii germinale intr-una din cele două alele ale genei INK4aassotsiirovany cu predispoziție ereditară la razvitiyumelanom piele (sindromul nev displazic). Unele dintre etihmutatsy inactiveaza doar de p16INK4a, fără a perturba funcția, prin urmare, pARF.V presupus că sindromul displazic nevusovsvyazan cu disfuncție este p16INK4a. Celulele nasledstvennyhi melanoamelor sporadice detectate modificări atât alleleygena INK4a, adică genă activă se comportă ca un opuholevyysupressor clasic. Cel mai adesea, al doilea (somatice) mutațiile predstavlyaetsoboy deleției genei INK4a duce la inactivarea p16INK4a ca, și PARF. Mutațiile, ștergerile și expresia metilarea INK4a genei vyzyvayuschiepoteryu de una sau ambele dintre produsele sale de proteine, nu numai în ereditar chastonablyudayutsya și melanoame sporadice, dar intr-un grup mare de alte tumori non-ereditare: cancer pancreatic, esofag, tractului biliar, vezică urinară și T B-celule de acute limfoblastice leucemie, mezoteliom, anaplasticheskihastrotsitomah, glioblastoame și altele.
3.5. Supresorul tumoral PTEN regleaza ciclul celular si apoptoza prin modularea căii de semnalizare PI3K-PKB / Akt
Pentru o supresoare tumorale, de multe ori greve in diverse tumori umane, și se referă gena PTEN. mutatii sale germinale provoca boala Cowden, care constă într-o predispoziție genetică la dezvoltarea hamartoame, cel mai adesea în glandele creierului, de san si tiroida. Inactivarea PTEN descrie, de asemenea, o parte semnificativă a diferitelor tumori non-ereditare - gliom, meningiom, melanomul, cancerul de rinichi, uter, san si prostata. Astfel, în timpul etapelor inițiale ale bolii este detectat, de regulă, se elimină numai una dintre alelele genei PTEN, in timp ce in tumori care sunt în etapele ulterioare de dezvoltare, de multe ori inactivate ambele alele. șoarecii transgenici cu knock-out a unei alele a PTEN gena marcat de dezvoltare într-o vârstă tânără diferite neoplasme: adenocarcinom intestinului, leucemie, limfosarcom si tumorile germinale produsului genei dr.Belkovy PTEN compus din 403 aminoacizi și are un grad ridicat de omologie cu fosfatazele dublă specificitate (defosforilat și tirozină și resturile de aminoacizi de serină / treonină). Caracteristica sa este abilitatea de a defosforila nu numai proteine, ci și lipide, în special fosfatidil-inozitolul (3,4,5) trifosfat (PIP-3) - un obiectiv important fosfatidil inozitid 3`-kinazei (PI3K), implicat în reglarea celulară ciclu și apoptoza. Este această activitate a PTEN proteine este responsabil, aparent, datorită funcției sale supresoare. Acest lucru este evidențiat în special prin faptul că cele mai multe mutatii gasite in celulele tumorale care sunt mapate în domeniul fosfatazei și anula defosforilare PIP-3, astfel inhibarea căii de semnalizare PI3K-PKB / Akt. Deoarece oncogene PKB / Akt inhiba calea mitocondriale de inducere a apoptozei prin mai multe mecanisme (inhibă Bad, activează Bcl2 etc.), pierderea funcției PTEN face ca celula mai puțin sensibilă la mulți stimuli apoptotice. In celule cu PTEN inactivat observat, de asemenea, stimularea proliferării celulare (expresie crescută a PKB / Akt crește nivelul ciclinei D). Restaurarea funcției PTEN în rezultatele celulelor tumorale, în funcție de context celular, sau apoptoza (cancer de prostată) sau oprirea ciclului celular în G1 (glioblastom, carcinom cu celule renale). Trebuie remarcat faptul că celulele gltoblastomy și cancerul de rinichi sunt astfel sensibilizate la acțiunea stimulov.PTEN apoptogenic suplimentare implicate, aparent, și în reglarea adeziunii și migrația celulară. Domeniul său omolog N-terminal Tenzin - proteine care interacționează în contactele focale cu citoscheletul actinei. Reziduuri Defosforiliruya tirozin kinazei FAK contactelor focale, PTEN pot inhiba formarea contactelor focale și celulă de împrăștiere precum și mișcarea lor.
 Anomalii ale genelor SOx și sindromul TVH Holt-Oram. Factori de creștere a fibroblastelor
Anomalii ale genelor SOx și sindromul TVH Holt-Oram. Factori de creștere a fibroblastelor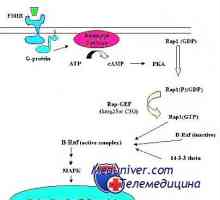 Mecanismul de acțiune al receptorilor gonadotropină. receptorii de epuizare
Mecanismul de acțiune al receptorilor gonadotropină. receptorii de epuizare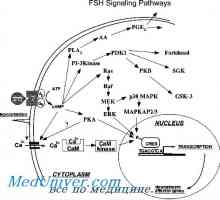 Gonadotropină receptori. Structura și funcția
Gonadotropină receptori. Structura și funcția Elongație mod și terminarea translației ARNm în timpul sintezei proteinelor
Elongație mod și terminarea translației ARNm în timpul sintezei proteinelor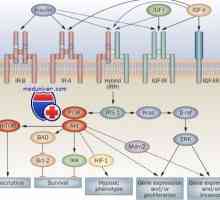 Receptorii cu activitatea tirozin kinazei. Receptorii pentru factorii de creștere și de insulină
Receptorii cu activitatea tirozin kinazei. Receptorii pentru factorii de creștere și de insulină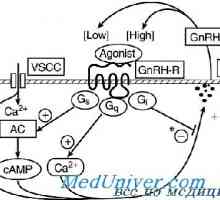 Receptor pentru a gonadotropinei de eliberare a hormonului. receptor Mutațiile GnRH
Receptor pentru a gonadotropinei de eliberare a hormonului. receptor Mutațiile GnRH Gene noi Descoperit responsabile pentru cancerul de san
Gene noi Descoperit responsabile pentru cancerul de san- Pătrundere în studiul de leucemie mieloidă acută
- Oamenii de știință au descoperit o gena care leaga cancerul de san si tiroidian
- Genele addutsina (ADD1, add2 și add3)
- Onkologiya-
- Onkologiya-
 Onkologiya-
Onkologiya- Onkologiya-
Onkologiya- Onkologiya-
Onkologiya-- Onkologiya-
 Onkologiya-
Onkologiya-- Onkologiya-
- Onkologiya-
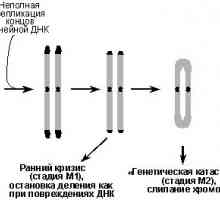 Onkologiya-
Onkologiya-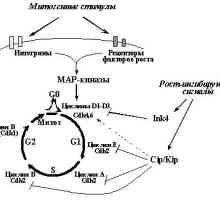 Onkologiya-
Onkologiya-
 Genetica cancerului
Genetica cancerului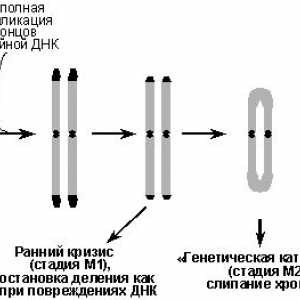 Onkologiya-
Onkologiya-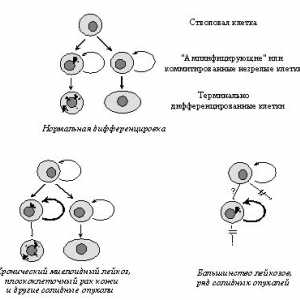 Onkologiya-
Onkologiya-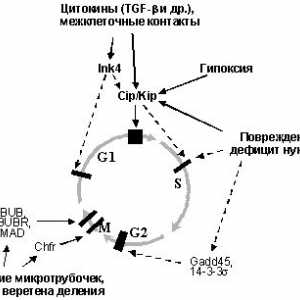 Onkologiya-
Onkologiya-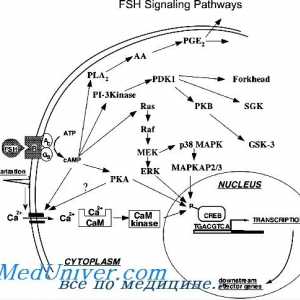 Gonadotropină receptori. Structura și funcția
Gonadotropină receptori. Structura și funcția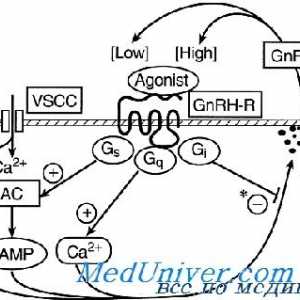 Receptor pentru a gonadotropinei de eliberare a hormonului. receptor Mutațiile GnRH
Receptor pentru a gonadotropinei de eliberare a hormonului. receptor Mutațiile GnRH